در افراد دارای کم خونی داسیشکل عمر گلبول قرمز کمتر است. در این افراد ترشح اریتروپویتین بالاتر است و در نتیجه همانندسازی در مغز استخوان بیشتر انجام میشود. مصرف فولیک اسید و B۱۲ در مغز استخوان بیشتر میشود.
ششمین آمینواسید در زنجیرهی بتای افراد دارای کمخونی داسی شکل به جای گلوتامیک اسید والین است.
در یک سلول ۶۱ کدون برای تولید پروتئین و ۳ کدون پایان وجود دارد.
کدون شروع همواره AUG است که آمینواسید متیونین جای آن مینشیند.
رنا از روی بخشی از یک رشته ساخته میشود.
ریبوزومها درون هسته حضور ندارند.
به ساخته شدن دنا از روی رنا رونویسی معکوس گفته میشود.
بیشترین رونویسی و ترجمه در G2 رخ میدهد.
رنابسپاراز به دو رشته متصل میشود.
در هسته سه نوع دنابسپاراز وجود دارد. در میتوکندری هم نوعی دنابسپاراز وجود دارد.
تنوع رنابسپاراز در یوکاریوتها بیشتر است، اما تنوع محصول رنابسپاراز در پروکاریوتها بیشتر است.
رنابسپاراز یک به طور مستقیم آنزیم تولید میکند.
رنابسپاراز سیتوپلاسمی کار هر سه رنابسپاراز را انجام میدهد.
متنوعترین رنا MRNA است.
رنابسپاراز دو و سه میتوانند رناهای دیگری هم تولید کنند.
جنس راه انداز دناییست.
در یوکاریوتها عوامل رونویسی به رنابسپاراز کمک میکند تا راه انداز را شناسایی کند و در پروکاریوتها وجود ندارد.
اتصال عوامل رونویسی به راهانداز قبل از شروع رونویسیست.
به اولین نوکلئوتیدی که دنابسپاراز روبهروی آن نوکلئوتید میگذارد جایگاه آغاز رونویسی نام دارد. درواقع جایگاه آغاز رونویسی یک نوکلئوتید است. پس راه انداز جایگاه آغاز رونویسی نیست.
دنابسپاراز یک نوع آنزیم است اما رنابسپارازها انواعی از آنزیمها هستند.
دو رشته دنا را رنابسپاراز باز میکند و توانایی شکستن پیوند هیدروژنی را دارد.
رنابسپاراز و هلیکاز به دو رشته دنا متصل میشوند اما دنابسپاراز به یک رشته متصل میشود.
حرکت رنابسپاراز همواره یک جهته است.
تک فسفاته کردن نوکلئوتید واکنشیست که توسط انواع آنزیمها انجام میشود مثل رنابسپاراز و دنابسپاراز.
ATP زمانی که در ساختار رنا قرار میگیرد مستقیما به AMP تبدیل میشود.
بعد از قرار گرفتن نوکلئوتید اول در جایگاه شروع رونویسی هیچ پیوند فسفودیاستری تشکیل نمیشود.
لزوما هر جفت نوکلئوتیدی که پیوند هیدروژنی آن توسط رنابسپاراز شکسته میشود مقابل آن نوکلئوتید گذاشته نمیشود.
راهانداز رونویسی نمیشود اما در همانندسازی شرکت میکند.
در مرحله آغاز رنابسپاراز به دنا میچسبد، دو رشته را باز میکند و یک زنجیره کوچک میسازد.
با جدا شدن بخشی از رنا از دنا ما وارد مرحله بعدی میشویم. درواقع در مرحله آغاز رنا از دنا جدا نمیشود.
در مرحله طویل شدن اتفاقات در سه محل بررسی میشود:
در پشت رنابسپاراز پیوند هیدروژنی بین ریبوز و دئوکسی شکسته میشود و بین دئوکسی و دئوکسی پیوند برقرار میشود. در جلوی رنابسپاراز پیوند هیدروژنی بین دئوکسی و دئوکسی شکسته میشود. در نقطهای که دنابسپاراز قرار دارد پیوند فسفودیاستر بین ریبوز و ریبوز تشکیل میشود. همچنین پیوند اشتراکی بین فسفاتهای ریبوز شکسته میشود. در این محل تشکیل هیدروژنی بین ریبوز و دئوکسی ریبوز هم دیده میشود.
بیشترینها (مثل بیشترین تشکیل فسفودیاستر) در مرحله طویل شدن رخ میدهد البته استثنائاتی هم دیده میشود. مثلا بیشترین تعداد نوکلئوتید دنا در مرحله پایان دیده میشود.
در هر سه مرحله رونویسی حرکت رنابسپاراز دیده میشود.
در مرحله رونویسی میتوان حرکت رنابسپاراز را بدون فعالیت بسپاراز مشاهده کرد.
به محض رسیدن به اولین نوکلئوتید توالی رونویسی مرحله پایان شروع میشود.
وقتی آخرین نوکلئوتید گذاشته شد ابتدا رنا میشود و سپس رنابسپاراز.
در رونویسی اولین پیوند تشکیل شده و آخرین پیوند تشکیل شده یکسان (هیدروژنی) هستند.
در مرحله آغاز رنا فقط درون حباب دیده میشود.
در باکتری ژنی وجود دارد که توالی آغاز دارد و توالی پایان ندارد.
در باکتری ژنی وجود دارد که نه توالی آغاز دارد و نه توالی پایان.
در باکتری ژنی وجود دارد که فقط توالی پایان دارد.
در یوکاریوتها:
اگر بین دو ژن یک راه انداز وجود داشته باشد همانندسازی هم جهت است.
اگر بین دو ژن یک توالی پایان باشد همانندسازی هم جهت است.
اگر بین دو ژن متوالی دو راه انداز باشد همانندسازی خلاف جهت است و رنابسپارازها از هم دور میشود.
اگربین دو ژن متوالی توالی پایان دیده نشود در یوکاریوتها مانند عبارت قبلی عمل میکنیم. اما توجه کنید که در پروکاریوتها قضیه میتواند مانند یوکاریوتها باشد یا اینکه پای قضیهی اپران وسط باشد. در قضیه اپران بین دو ژن توالی پایان دیده نمیشود و رونویسی همجهت است.
اگر بین دو ژن متوالی دو توالی پایان دیده میشود همانندسازی مخالف هم است و رنابسپارازها به هم نزدیک میشوند.
اگر بین دو ژن متوالی راه انداز دیده نمیشود قضیه میتواند مثل عبارت قبلی باشد. اما توجه کنید استثنایی مانند اپران وجود دارد.
پروتئینسازی هموگلوبین در دوران جنینی انجام میشود.
سلول در جی یک باید رشد زیادی را تجربه کند و ساخت درRRNA در این مدت بسیار زیاد است.
رنا پس از ساخته شدن تغییراتی انجام میدهد. یکی از این تغییرات پیرایش است. پیرایش در باکتری وجود ندارد.
رنای پیک ممکن است حین رونویسی تغییر کند. پس از رونویسی هم ممکن است تغییراتی در رنای پیک ایجاد شود. یکی از این تغییرات پیرایش است.
MRNAای که از منافذ غشا عبور میکند حتما پیرایش شده است.
دو سر MRNA همواره اگزون است. تعداد اگزونها از اینترونها بیشتر است.
در پیرایش تشکیل فسفودیاستر از شکستن آن کمتر است.
همواره تعداد نوکلئوتید رنا از ژن سازنده آن کمتر است. دقت کنید ژن دو رشتهایست.
ویژگیهای منحصربهفرد هر آمینواسید به گروه R بستگی دارد. دقت کنید همهی ویژگیهای آمینواسیدها به گروه R بستگی ندارد. آمینواسید ویژگیهای اسیدی و آمینی هم دارد که به گروه اسید و آمین وابسته است.
در ساختار پروتئینها حداکثر بیست نوع آمینواسید میتوان دید.
نوع، ترتیب و تعداد آمینواسیدها در پروتئینها عمل آنها را تعیین میکند.
هر آمینواسید میتواند در شکلدهی پروتئین موثر باشد و تاثیر آن به ماهیت گروه R بستگی دارد.
شکل پروتئینها عمل آنها را مشخص میکند.
تنها آنزیم غیرپروتئینی بدن rRNA است.
رشتههای پروتئینی همواره خطی و بدون انشعاب هستند.
آمینواسید شماره یک از انتهای آمینی شروع میشود.
آمینو اسید اول و آمینواسید آخر در یک پیوند پپتیدی شرکت میکنند. بقیه آمینواسیدها دو پیوند پپتیدی میدهند.
هر تغییری PH منجر به تغییر شکل پروتئين نمیشود.
تغییر در پیوندهای پروتئينها میتواند منجر به تغییر شکل آن بشود.
اولین پروتئينی که ساختار آن مشخص شد میوگلوبین بود. دقت کنید میوگلوبین اولین پروتئين شناسایی شده نبود.
در سطح اولین همهی آمینواسیدها در پیوندهای پپتیدی شرکت میکنند.
در سطح دوم بعضی از آمینواسیدها در پیوند هیدروژنی شرکت میکنند. این پیوند بین گروه هیدروژن یکی از آمینها و اکسیژن برقرار میشود.
در سطح دوم پروتئينها چهار شکل وجود دارد:
۱-فقط مارپیچی ۲-فقط صفحهای ۳-مارپیچی و صفحهای ۴-ساختارهای دیگر که کتاب درسی در مورد آنها نمیگوید.
آمینواسیدها براساس گروه R به آمینواسیدهای آبدوست و آبگریز تقسیم میشوند. این موضوع مبنای تشکیل ساختار سوم است که در آن پروتئينها ساختار مچاله و نامتقارن به خود میگیرند.
تا خوردگی پروتئین از سطح دو آغاز میشود. تا خوردگی بیشتر در سطح سوم وجود دارد.
علت تشکیل ساختار اول پیوندهای پپتیدیست. علت تشکیل ساختار دوم پیوندهای هیدروژنیست. علت تشکیل ساختار سوم برهمکنشهای آبگریز است. در ساختار سوم پیوندهای هیدروژنی، اشتراکی و یونی هم تشکیل میشود که به خاطر تثبیت ساختار سوم است.
در سطح یک فقط پیوند پپتیدی دیده نمیشود. در سطح یک پیوندهای درون هر آمینواسید هم دیده میشود. دقت کنید بین دیده شدن و تشکیل شدن تفاوت وجود دارد. در سطح یک تنها پیوند پپتیدی تشکیل میشود، اما پیوندهای دیگری هم وجود دارند.
میوگلوبین نمونهای از یک پروتئین با ساختار سوم است.
زیرواحد آلفا و بتای هموگلوبین هرکدام به تنهایی ساختار سوم را دارند. این زیرواحدها در سطح دوم به شکل مارپیچ درآمدهاند.
در ساختار مارپیچ پیوند هیدروژنی بیشتری برقرار میشود.
در ساختار مارپیچ مولکولهای R خارج از مارپیچ قرار میگیرند.
همهی آنزیمهای ترشحی برهمکنش آبگریز دارند.
در سطح یک، دو و سه فقط یک آمین آزاد دیده میشود.
شروع تشکیل مرحله اول بر شروع تشکیل مرحله دوم مقدم است. همچنین ایجاد مرحله دوم زودتر از ایجاد مرحله سوم رخ میدهد، اما دقت کنید که شروع مرحله دوم میتواند قبل از اتمام تشکیل مرحله اول شروع شود. اما تشکیل سطح چهارم نمیتواند همزمان با مراحل یک، دو یا سه رخ بدهد.
در مرحله یک پیوند هیدروژنی وجود ندارد.
اکتین و میوزین در انقباض هر سه نوع ماهیچه دخیل هستند.
واکنشهای شیمیایی در صورتی انجام میشود که انرژی اولیه کافی برای انجام آزمایش فراهم باشد.
آنزیمها امکان برخورد مناسب مولکولها را افزایش و انرژی فعالسازی را کاهش میدهد.
آنزیمهای بدن نمیتوانند سرعت همهی واکنشها را زیاد کند.
بدون حضور آنزیم در دمای بدن سوخت و ساز سلولها بسیار کند انجام میشود (نه اینکه اصلا انجام نشود).
چند نکته در مورد آنزیمها:
آیا هر آنزیمی که در سلول وجود دارد توسط آن سلول ساخته شده است؟ نه. مثلا آنزیمهای القای مرگ برنامهریزی میتواند توسط یک سلول دیگر وارد شده باشد.
آیا هر آنزیمی درون سلول ساخته میشود؟ نه. مثلا پپسین درون معده ساخته میشود.
آنزیمهای اسپرم در بدن مرد ساخته میشود و در بدن زن فعالیت میکند.
برای ورود آنزیمها (یا پروتئینهای درشت) الزاما از درونبری استفاده نمیشود. مثلا آنزیم مرگ برنامهریزی شده توسط سلول کشنده طبیعی از طریق سوراخی که پرفورین ایجاد میکند وارد سلول میشود.
rRNA هم آزیم است و انرژی فعالسازی را کاهش میدهد. همهی آنزیمها انرژی فعالسازی را کاهش میدهند.
آنزیمها در ساختار خود جایگاه فعال دارند که اختصاصی عمل میکنند. فقط مولکولهایی میتوانند در جایگاه فعال بنشینند که با آن مکمل (نه مشابه) باشند.
بعضی (نه همه) آنزیمها برای فعالیت خود به یونهای فلزی مانند آهن و مس یا مواد آلی مثل ویتامینها نیاز دارند.
سوال: آیا هر پروتئینی که برای کار خود نیاز به اتم آهن دارد لزوما آنزیم است؟ نه. مثلا هموگلوبین در ساختار خود اتم آهن دارد.
به مواد آلی (مثل ویتامینها) که به آنزیمها کمک میکند کوآنزیم میگویند. دقت کنید به مواد معدنی (مثل اتم آهن) کوفاکتور گفته میشود.
در همه کوآنزیمها اتم کربن وجود دارد.
بعضی از مواد سمی میتواند در جایگاه فعال آنزیم قرار بگیرد و مانع فعالیت آن بشود، اما بعضی از مواد سمی پیشماده آنزیمها هستند. مثلا برخی آنزیمهای کبدی میتواند آمونیاک را به اوره تبدیل کند. درواقع در کبد مواد سمی مصرف میشوند.
آنزیمها عمل اختصاصی دارند و روی یک یا چند پیش ماده خاص اثر دارند.
آنزیمها لزوما در همه واکنشهای بدن شرکت نمیکنند. مثلا اولین تبدیل پپسینوژن به پپسین توسط HCL شرکت میکنند.
لزوما افزایش یا کاهش PH روی عملکرد پروتئين موثر نیست. چیزی که عملکرد پروتئين را مختل میکند تغییر PH در حدیست که از PH بهینه خارج شود.
دنابسپاراز در بیضه در دمای ۳۴ درجه فعالیت میکند. همین آنزیم در دیگر قسمتهای بدن در دمای ۳۷ درجه فعالیت میکند.
بیشترین فعالیت آنزیم با بهترین فعالیت آنزیم متفاوت است. بهترین فعالیت در دمای بهینه انجام میشود. بیشترین فعالیت در یک گستره دمایی خاص انجام میشود.
آنزیمها در دمای بالاتر غیرفعال میشوند که این غیرفعال شدن دائمی و برگشتپذیر است. در دمای پایینتر هم آنزیمها غیرفعال میشوند، منتهی این غیرفعال شدن برگشتپذیر است.
در طرح نیمه حفاظتی و غیرحفاظتی ما تشکیل پیوند هیدروژنی بین نوکلئوتیدهای جدید و قدیمی را داریم.
در طرح نیمه حفاظتی ما بین رشته قدیم و رشته جدید پیوند هیدروژنی داریم.
در طرح نیمه حفاظتی شکستن پیوند فسفودیاستر قطعا دیده میشود.
در آزمایش مزلسون و استال با گذشت هر دور همانندسازی ضخامت نوار بالایی بیشتر میشود.
در همانندسازی نوکلئوتید ساخته نمیشود. ما در طی همانندسازی یک رشته DNA جدید خلق میکنیم.
هلیکاز نقاط شروع همانندسازی را شناسایی میکند، مارپیچ DNA را باز میکند و سپس دو رشته را از هم جدا میکند.
در هر حباب همانندسازی بیش از شش آنزیم دیده میشود.
هلیکاز برای شروع فعالیت خود به دو رشته DNA متصل میشود. اما DNAپلیمراز برای شروع کار خود به یک رشته متصل میشود.
دنابسپاراز توانایی شکستن پیوند فسفات را دارد. توانایی تشکیل پیوند کوالانسی بین نوکلئوتیدها را هم دارد. دنابسپاراز توانایی شکستن پیوند فسفودیاستر بین نوکلئوتیدها را هم دارد (در فرایند ویرایش).
هلیکاز همواره در یک جهت کار میکند. دنابسپاراز معمولا در یک جهت فعالیت میکند، اما در فرایند ویرایش توانایی برگشت هم دارد.
اشتباه اصلاح نشده دنابسپاراز جهش نام دارد. این اشتباه در نقطه وارسی G1 بررسی میشود.
تشکیل پیوند هیدروژنی بر تشکیل پیوند فسفودیاستر مقدم است، اما در زمانی که دنابسپاراز اشتباه میکند پیوند هیدروژنی بین نوکلئوتیدهای متقابل تشکیل نمیشود و تشکیل پیوند فسفودیاستر بر پیوند هیدروژنی مقدم است (در درستی این گفته شک دارم).
امکان خطا در دنابسپاراز را وسیع درنظر بگیرید. یعنی روبهروی نوکلئوتید A همهی انواع نوکلئوتیدها حتی خود A هم قرار میگیرد.
برگشتن دنابسپاراز بعد از ایجاد هر پیوند فسفودیاستر رخ میدهد. حالا این برگشتن و بررسی ممکن است به تغییر نوکلئوتید منجر شود.
قبل از همانندسازی دنا چند کار انجام میشود :
۱-پیچ و تاپ فامینه باز میشود (پیچ و تاب فامینه از مارپیچ دنا متفاوت است). ۲-پروتئینهای همراه DNA جدا میشود (هیستون مهمترین پروتئین همراه DNA است).
دو کار بالا توسط آنزیمهایی انجام میشود:
الف-این آنزیمها بیشتر از یکی هستند. ب-هلیکاز و دنابسپاراز جز این آنزیمها نیستند.
در همانندسازی DNA هم پروتئینهای همراه DNA جدا میشود اما به یک موضوع توجه کنید:
جدا شدن هیستونها در DNA خطی دیده میشود و در DNA حلقوی اصلا هیستون وجود ندارد.
حالا همانندسازی شروع میشود:
DNA خطی قطعا چند جایگاهی و قطعا دو جهته است. در DNAهای خطی همواره تعداد جایگاه پایان بیشتر از جایگاه آغاز است.
همانندسازی در DNA حلقوی چهار حالت دارد:
۱-یک جایگاه همانندسازی و یک جهته. ۲-یک جایگاه همانندسازی و دو جهته. ۳-چند جایگاه همانندسازی و یک جهته. ۴-چند جایگاه همانندسازی و دو جهته.
در تمام حالتهای همانندسازی DNA حلقوی تعداد جایگاه آغاز و تعداد جایگاه پایان با هم برابر است.
نکته: سلولهایی که در G0 قرار دارند اصلا جایگاه آغاز و پایان تشکیل نمیدهند.
اگر دنابسپاراز در سیتوپلاسم فعالیت کند تعداد نوکلئوتیدهای داخل هسته تغییر نمیکند.
در حباب همانندسازی نوکلئوتید با باز آلی U دیده میشود، اما استفاده نمیشود.
در حباب همانندسازی دو نوع پیوند هیدروژنی دیده میشود:
۱-پیوند هیدروژنی بین رشته قدیم و رشته جدید ۲-پیوند هیدروژنی بین رشتههای قدیمی
هلیکاز پیوند هیدروژنی از نوع دوم را میشکند و به نوع اول کاری ندارد.
باکتری یک DNA حلقوی متصل به غشا دارد، اما ممکن است این DNA تنها DNA باکتری نباشد. معمولا باکتریها DNAهای کمکی به نام پلازمید دارند. این DNAها میتواند یکی یا بسیار بیشتر باشد.
ژن مقاومت به آنتیبیوتیک روی DNA اصلی باکتری قرار ندارد.
در باکتری حین همانندسازی میتوان DNA خطی را در باکتری دید.
در یوکاریوتها دنا در هر فامتن به صورت خطیست. پس به DNA حلقوی میتوکندری و پلاستها فامتن نمیگوییم.
فقط در یوکاریوت و در DNA خطی یوکاریوت تعداد جایگاه آغاز همانندسازی بسته به مراحل رشد و نمو تغییر میکند.
تعداد جایگاههای آغاز همانندسازی در باکتری میتواند تغییر کند. درواقع با افزایش پلازمیدها جایگاههای آغاز همانندسازی در باکتری افزایش پیدا میکند، اما همچنان تعداد جایگاه آغاز در DNA اصلی بدون تغییر مانده است.
تعداد جایگاه آغاز در سلولهایی که سرعت تقسیم بیشتری دارند (مثل سلولهای مغز استخوان) میتواند از بقیه سلولها بیشتر باشد.
تعداد جایگاه همانندسازی در کروموزومهای مختلف یک سلول هم متفاوت است. مثلا اندازه کروموزوم شماره یک و ایگرگ در مردان را با هم مقایسه کنید. فکر میکنید جایگاه همانندسازی کدام بیشتر است؟
سرعت همانندسازی در جایگاههای مختلف یک دنا هم میتواند متفاوت باشد.
در جاهایی از دنا که نوکلئوتید C و G بیشتری وجود دارد سرعت هلیکاز کمتر و مصرف انرژی بیشتر است.
کتاب درسی مقدمهای آورده که بررسی آن میتواند یک یادآوری خوب از گذشته باشد.
هر یک از سلولهای بدن ما ویژگیهایی مانند شکل و اندازه دارد. این ویژگیها تحت فرمان هسته است.
ویژگیهایی مانند شکل و اندازه تحت فرمان کروموزومها هستند. با کروموزومها در سال یازدهم آشنا شدیم.
کروموزومها میتوانند در هسته حضور داشته باشند یا در سیتوپلاسم و متصل به غشا باشند. در جانداران یوکاریوتی مثل انسانها کروموزوم در هسته حضور دارد و در بعضی از جانداران مثل پروکایوتها کروموزومها در هسته نیستند چون هسته وجود ندارد.
علاوه بر سلولهای پروکاریوتی برخی از سلولهای یوکاریوتی هم میتوانند بدون هسته باشند، مانند گلبولهای قرمز.
دستورالعملهای هسته در حین تقسیم از سلولی به سلول دیگر و در حین تولید مثل از نسلی به نسل دیگر منتقل میشود.
بد نیست یک بار دیگر مرور کنیم: همهی سلولهای بدن ما هستهدار نیستند.
سال گذشته با فرایند تقسیم آشنا شدیم و میدانیم همهی سلولهای بدن تقسیم نمیشوند. گلبولهای قرمز، اسپرمها و یا سلولهای پادتنساز در قرار دارند و تقسیم نمیشوند.
سلولها، هستهدار یا بدون هسته اگر فرایند تقسیم را انجام بدهند دستورالعملهای خود را به سلول بعد منتقل میکنند. به جز تقسیم، مفهوم دیگر هم به نام تولید مثل وجود دارد. اگر تولید مثل انجام شود DNA به نسل بعد منتقل میشود.
همهی جانداران تولید مثل انجام نمیدهند. مثلا برخی از زنبورهای عسل با اینکه جاندار و موجود زنده محسوب میشوند اما تولید مثل انجام نمیدهند.
آزمایش گریفیت
باکتری کپسولدار سلول را آلوده و بیمار میکند. باکتری بدون کپسولدار سلول را آلوده میکند، اما منجر به بیماری نمیشود.
در هر چهار مرحله به دلیل تزریق از ماستوسیتهای آسیب دیده هیستامین رها شده و موجب التهاب میشود.
در هر چهار مرحله آزمایش گریفیت ما ترشح پادتن را داریم.
در مرحله یک و چهار باکتری کپسول میسازد.
گریفیت نمیدانست DNA چیست یا چطور منتقل میشود، اما یک نتیجه بسیار مهم گرفت: ماده وراثتی میتواند انتقال یابد.
از آزمایش گریفیت میتوان نتیجه گرفت که DNA و پوشینه برخلاف غشای سلول نسبت به گرما مقاوم است.
همه باکتریهای بدون کپسول، کپسولدار نمیشوند. فقط برخی از آنها این ویژگی را کسب میکنند.
باکتری نومونیای بدون پوشینه و پوشینه دار از یک گونه هستند. این یعنی با انتقال ژن از باکتری پوشینهدار به باکتری بدون پوشینه جاندار تراژن تولید نمیشود.
عامل آنفولانزا ویروس است و عامل سینه پهلو میکروب.
گریفیت روی موش و باکتری آزمایش کرد. موش یک جانور است و باکتری یک جاندار.
آزمایش ایوری
ایوری در مرحله اول و آخر از آنزیم استفاده کرد اما در مرحله دوم خبری از آنزیم نبود.
ایوری در مرحله اول فقط از یک آنزیم استفاده کرد، اما در مرحله آخر از انواعی از آنزیمها استفاده کرد.
در مرحله دو و سه عصاره به چند قسمت تقسیم شد. این تقسیم شدن در مرحله دو بر اساس چگالی و در مرحله آخر صرفا به بخشهای مختلفی تقسیم شد.
ایوری روی باکتری آزمایش کرد که یک جاندار است نه یک جانور.
در آزمایش اول ایوری عصاره باکتری را به داخل لوله آزمایش میریزد و پروتئينهای داخل آن را تجزیه میکند. چند نکته در رابطه با این موضوع وجود دارد: -اولا خود آنزیم نوعی پروتئين است. درست است که در لوله آزمایش دیگر پروتئين عصاره باکتری دیده نمیشود، اما آنزیم وجود دارد که خود نوعی پروتئین است. -حاصل تجزیه پروتئينها آمینواسید است. پس بعد از تجزیه پروتئين در لوله آزمایش آمینواسید دیده میشود.
در آزمایش آخر مرحلهای وجود دارد که ایوری به کمک آنزیم کربوهیدراتها را تجزیه میکند. دقت کنید با اینکه کربوهیدراتها همگی تجزیه میشوند اما همچنان درون دنا، رنا و نوکلئیکاسیدهای سلول کربوهیدرات وجود دارد.
در مرحله دوم آزمایش ایوری هر کدام از مولکولهای زیستی به تنهایی وارد محیط کشت میشوند.
نوکلئیک اسیدها
نوکلئوتیدها از چند چیز تشکیل شدهاند:
۱-قند که در RNA ریبوز و در DNA داکسی ریبوز است. ۲-فسفات. نوکلئوتیدهایی که در سلول آزاد هستند سه فسفات دارند و نوکلئوتیدهایی که در زنجیره مینشینند یک فسفات دارند. ۳-باز آلی نیتروژندار که چهار مدل است؛ A،T،C،G. بعدها مشخص شد که در RNAها به جای باز آلی T باز آلی U وجود دارد. دقت کنید که در DNA باز آلی U وجود ندارد.
نوکلئوتیدها بر اساس حلقه باز آلی دو دسته هستند: پورین و پیریمیدین.
باز آلی نوکلئوتیدهای A و G دو حلقهای است. درواقع «آق (AG) پورین دو حلقه دارد».
باز آلی نوکلئوتیدهای T و C و U تک حلقهای است که به آنها پیریمیدین میگویند.
گروه فسفات با پیوند اشتراکی (کووالانسی) به قند پنج کربنه متصل شده است. باز آلی نیتروژندار هم با پیوند اشتراکی به قند پنج کربنه وصل شده است. دقت کنید گروه فسفات و باز آلی نیتروژندار با هم پیوند ندارند.
در نوکلئوتیدهای پورینی قند پنج کربنه به پنج ضلعی باز آلی متصل شده (این نوکلئوتیدها بازها دو حلقهای هستند).
در نوکلئوتیدهای پیریمیدینی قند به باز آلی شش ضلعی متصل است (باز آلی در این نوکلئوتیدها تک حلقهایست).
قند در نوکلئوتیدها پنج کربنه است، اما حلقهی این قند چهار کربن دارد. یکی از کربنهای این نوکلئوتید در خارج از حلقه قرار دارد.
ATP نوعی نوکلئوتید با باز آلی آدنین و قند ریبوز است. این مولکول نه در ساخت DNA شرکت میکند و نه در ساخت RNA.
ما در هر نوکلئوتید حلقه شش ضلعی میبینیم و صد در صد در هر نوکلئوتید فقط یک حلقه شش ضلعی میبینیم.
به فاصله قند تا قند پیوند فسفودیاستر میگویند.
در نوکلئوتیدها پیوند فسفودیاستر دیده نمیشود، اما آنها مانند رناها و دناها پیوند قند فسفات دارند.
بین RNAها و DNAها هیچ نوکلئوتید مشترکی وجود ندارد.
یک سر رنا فسفات دارد و سر دیگر هیدروکسیلدار است.
مولکول دنا دو سر یکسان دارد. اما هر رشتهی دنا دو سر متفاوت دارد.
جهتگیری قندها در دو رشته یک دنا ناهمسو است.
رنای حلقوی در میتوکندری در مجاورت ریبوزومها قرار میگیرد.
اگر نوکلئیک اسید فرضی فقط دارای باز پورین یا فقط دارای باز پیریمیدین باشد آن نوکلئيک اسید فقط از جنس RNA است.
آزمایش چارگاف
چارگاف متوجه شد که تعداد نوکلئوتیدهایی که دارای باز آلی A هستند با نوکلئوتیدهایی که دارای باز آلی C هستند برابر است. این موضوع در مورد باز C و G هم صادق است.
چارگاف متوجه نشد که نوکلئوتیدها مقابل هم مینشینند. او حرفی از بازهای مکمل نزد. او صرفا متوجه شد چه نوکلئوتیدهایی در سلول با هم برابر هستند.
در یک مولکول DNA تعداد نوکلئوتیدهای پورینی و پیریمیدینی با هم برابر است. یعنی در یک مولکول DNA (نه در یک رشته) نصف تعداد نوکلئوتیدها پورینی و نصف دیگر پیریمیدینی هستند.
ویلکینز و فرانکلین
اولین کسانی که مشخص کردند DNA مارپیچیست ویلکینز و فرانکلین بودند، اما واتسون کریک اولین کسانی بودند که متوجه شدند DNA دورشتهایست.
واتسون و کریک
واتسون و کریک مشخص کردند که مارپیچ DNA دو رشتهایست. آنها فهمیدند که نوکلئوتیدهای C با G و نوکلئوتیدهای A با T جفت میشود.
همواره در برابر یک نوکلئوتید پورینی یک نوکلئوتید پیریمیدینی مینشیند. این موضوع باعث میشود همواره در برابر یک باز یک حلقه، یک باز دو حلقهای بنشیند. این کار باعث میشود که قطر DNA درسراسر آن ثابت بماند و همین موضوع باعث پایداری مولکول DNA میشود.
مولکول DNA فرضی با بیشترین پایداری: مولکول که در آن فقط C و G وجود دارد.
مولکول DNA فرضی با بیشترین پایداری: مولکول که در آن فقط A و T وجود دارد.
در نومونیا کپسول با غشا در تماس نیست و دیواره باکتری مانع از تماس کپسول و غشا میشود.
در DNA خطی تعداد نوکلئوتید از تعداد فسفودیاستر بیشتر است.
در DNA حلقوی تعداد نوکلئوتید برابر با تعداد فسفودیاستر است.
تعداد پیوند فسفواستر دو برابر تعداد پیوند فسفودیاستر در DNA حقلوی است. در DNA خطی تعداد فسفواستر از فسفودیاستر بیشتر است اما دوبرابر نیست.
در DNA حلقوی هر قند به دو فسفات و هر فسفات به دو قند متصل است. در DNA خطی این موضوع صادق نیست، مگر اینکه میانه DNA را در نظر بگیریم.
DNA چه حلقوی باشد چه خطی، همواره مارپیچیست.
در پلهها هم پیوند هیدروژنی و هم پیوند اشتراکی دیده میشود. در ستونها فقط پیوند اشتراکی دیده میشود.
رنا و انواع آن
ماده وراثتی با ماده حاوی اطلاعات وراثتی متفاوت است. ماده وراثتی همان DNA است. ماده حاوی اطلاعات وراثتی میتواند DNA یا RNA باشد.
اگر در مورد نوکلئیک اسیدها سوال شد منظور هم DNA است و هم RNA.
اگر در مورد نوکلئیک اسید خطی سوال شد هم DNA خطیست و هم بیشتر RNAها.
ما نوکلئيک اسید خطی در باکتری داریم، اما DNA خطی در باکتری وجود ندارد.
RNA میتواند مارپیچی یا خطی باشد.
در ساختار RNAهایی مانند TRNA میتوان پیوند هیدروژنی را دید.
در جریان تقسیم هم DNA و RNA به نسل بعد منتقل میشود.
در تولید مثل جنسی لزوما ما لقاح نداریم. مثلا در زنبور بکرزایی وجود دارد که گامت ماده شروع به تقسیم میکند و زنبور نر را به وجود میآورد.
لقاحها دو دسته هستند، داخلی و خارجی.
در لقاح داخلی لزوما دو نفر شرکت ندارند و جانورانی هستند که هم گامت ماده و گامت نر را تولید میکنند.
در لقاح داخلی گامت از یک نفر خارج و وارد نفر دوم میشود. در اکثر موارد شخص گامت دهنده نر است، اما برخی موارد وجود دارد که شخص دهنده گامت ماده است (مثل اسبک ماهی).
هر جانوری که رحم دارد لزوما پستاندار نیست. مثلا کرم کبد رحم دارد.
هر پستانداری هم لزوما رحم ندارد. مثلا پلاتی پوس رحم ندارد و رحم در جانوران از کانگرو شروع میشود که رحم ابتدایی دارد.
گامت سالم در همه جانوران هاپلوئید است.
در همه جانوران اسپرم دارای تاژک و دارای حرکت و تخمک فاقد حرکت است.
در همه جانوران تخمک دارای اندوخته غذاییست.
همه حشرات لقاح داخلی دارند.
هر جانوری که لقاح داخلی دارد دارای دستگاه تولید مثل با اندامهای تخصصیافته است.
تولید مثل در مهرهداران
-ماهیها: بیشتر لقاح داخلی و برخی مانند اسبکماهی لقاح خارجی دارند. -دوزیستان: تماما لقاح خارجی دارند. -خزندگان، پرندگان و پستانداران لقاح داخلی دارند.
این موجودات قطعا دارای دستگاه تولید مثل با اندامهای تخصصیافته هستند.
اندوخته غذایی جنین:
-در ماهیها و دوزیستان به دلیل دوره جنینی کم ذخیره غذایی تخمک کم است. -در پرندگان و خزندگان به دلیل تخمگذاری ذخیره غذایی تخمک کم است. -در پستانداران ذخبره تخمک به دلیل ارتباط خونی جنین با مادر کم است، البته با یک استثنا: پلاتیپوس تخمگذار است و ذخیره غذایی آن زیاد است.
در کرم کبد دمای کیسه بیضه و تخمدان تقریبا برابر است.
در کرم کبد هر چه به سمت انتها حرکت میکنیم بدن ضخیمتر میشود.
کرم کبد تولید مثل جنسی یک والدی با لقاح دارد.
زنبور ملکه و مار میتوانند تولید مثل جنسی یک والدی بدون لقاح دارد.
محتوای ژنتیکی بچهی مار حاصل از بکرزایی با محتوای ژنتیکی مادر یکسان نیست.
گامت زنبور ماده میتواند تقسیم شود. گامت بقیه جانوران در G0 میماند.
در یکی از شکلهای کتاب لوله فالوپ کلا از رحم پایینتر است. در شکلهای دیگر بخشی از لوله فالوپ بالاتر از رحم قرار میگیرد.
تخمدان تخممرغی شکل است. بخش نازک آن توسط طناب پیوندی عضلانی به بخش خارجی رحم متصل است.
بخش پیوندی طناب پیوندی عضلانی به تخمدان و بخش عضلانی آن به دیوار خارجی رحم متصل است.
حرکت زوائد انگشت مانند باعث ورود اووسیت ثانویه به لوله رحم میشود. زوائد انگشت مانند دارای ماهیچه صاف هستند و دستگاه عصبی خودمختار آنها را کنترل میکند.
قسمت پهن تخمدان به زوائد انگشتی نچسبیده است.
در زنان گامت در غدد جنسی تولید نمیشود. درواقع سلولی که وارد لوله فالوپ میشود اووسیت ثانویه است نه گامت.
زنش مژکهای لوله فالوپ از ته به سر لوله است. مسیر حرکت اسپرم مخالف زنش مژکها و مسیر حرکت اووسیت ثانویه (نه گامت) موافق زنش مژکهاست.
اسپرمها به وسیله حرکت تاژک در لوله فالوپ حرکت میکنند. اووسیت ثانویه توسط زنش مژکهای لوله فالوپ حرکت میکند.
قطر رحم، با قطر دیواره رحم فرق میکند. قطر رحم هر چه به سمت پایین حرکت میکند کم میشود. درواقع بخش پهن رحم بخش بالایی آن میشود. دیواره رحم هر چه به سمت گردن آن میرود ضخیمتر میشود.
سقف رحم و گردن رحم دیواره ضخیمی دارد.
رحم مژک ندارد. لوله فالوپ مژک دارد است. دیواره داخلی رحم چین خورده است اما مژک ندارد.
تخمدان درون حفره شکمی قرار دارد. پرده صفاق با تخمدانها در تماس است.
دوره جنسی زنان با بلوغ آغاز میشود، اما گامتزایی آنها در دوره جنینی رخ میدهد.
به مجموع اووسیت و و سلولهای تغذیه کننده آن فولیکول میگویند.
بعد از تولد تعداد فولیکول ثابت میماند، اما تعداد سلولهای تغذیه کننده اووسیت افزایش مییابد.
رحم مانند مثانه اندامی کیسهای شکل است.
فشار روحی میتواند باعث سریعتر شدن روند از کار افتادن تخمدانها شود. پس میتوان گفت افزایش هورمون کورتیزول هم میتواند باعث این موضوع شود.
زمانی که فولیکول هنوز کوچک است اووسیت اولیه در مرکز آن قرار دارد. با رشد فولیکول و پیدا شدن حفره فولیکولی اووسیت اولیه به حاشیه رانده میشود.
در زمان تخمکگذاری چهار چیز از فولیکول خارج میشود: اووسیت ثانویه، اولین جسم قطبی، سلولهای تغذیه کننده، مایع فولیکولی.
تنها فولیکول بالغ به دیواره تخمدان چسبیده است.
لایه ژلهای شفاف دور اووسیت درون تخمدان تشکیل میشود.
FSH باعث تقسیم سلولهای تغذیه کننده فولیکولی میشود، اما باعث حجیم شدن آنها نمیشود. چیزی که در فولیکول حجیم میشود لایههای این سلولهاست نه خود سلولها.
سلول لقاح دهنده:
-از طرف مردها حتما یک گامت است (اسپرم). -از طرف زنها میتواند یک گامت (تخمک) یا جسم قطبی باشد.
سلول حاصل از لقاح میتواند تقسیم شود و یک توده ایجاد کند و سپس دفع شود (سلول حاصل از لقاح اسپرم و جسم قطبی).
افزایش LH عامل اصلی تخمکگذاریست.
LH و FSH روی رحم گیرنده ندارد.
ما در ابتدا چرخه رحمی کاهش ضخامت رحم و در ادامه افزایش ضخامت رحم را شاهد هستیم.
اگر بارداری ندهد در پایان مرحله لوتئالی ما کاهش ضخامت رحم را شاهد خواهیم بود.
هر کاهش ضخامت آندومتر قاعدگی نیست.
در مرحله لوتئال ما خونریزی نداریم.
ماهیچه صاف رحم در قاعدگی تخریب نمیشود.
در سطحی در اندومتر که در نزدیکی ماهیچه صاف قرار دارد پیچخوردگی سرخرگ از سیاهرگ بیشتر است.
سرخرگهای آندومتر از سیاهرگهای آن بیشتر است.
پروژسترون در افزایش ترشحات دیواره رحم اثرگذار است.
دستگاه تولید مثل در مرد فقط یک هورمون تولید میکند. این دستگاه در زنان دو هورمون تولید میکند که میشود هورمونها.
برای تمایز صحیح اسپرمها چه چیزی نیاز است؟
۱-دما ۲-ترشحات سرتولی ۳-هورمون FSH.
جریان خون دمای بیضه را با بقیه بدن یکسان نمیکند. در بیضه شبکهای از رگهای کوچک دمای آن را صرفا تنظیم میکند.
در دستگاه تناسلی ما سه نوع مجرا و لوله میبینیم:
۱-مجرای اسپرمساز ۲-اپیدیدیم ۳-مجرای اسپرمبر
مجرای اسپرمساز متعدد است. اپیدیدیم در هر مرد دوتاست. مجرای اسپرمبر هم دو تا داریم.
هر سه مجرا، لوله یا لولههایی پیچیده هستند. پیچیده بودن مجرای اسپرمبر در سراسر آن یکسان نیست.
دمای اسپرمساز و اپیدیدیم ۳۴ درجه است. مجرای اسپرمبر در بخشی ۳۴ درجه و در بخشی دیگر ۳۷ درجه است.
مجرای اسپرمساز درون بیضه قرار دارد. اپیدیدیم خارج از بیضه است اما درون کیسه بیضه قرار دارد. بخشی از مجرای اسپرمبر درون کیسه بیضه و بخشی از آن درون محوطه شکم قرار دارد.
اسپرم موجود در مجرای اسپرمساز توانایی حرکت ندارد. اسپرمهای درون اپیدیدیم دو مدل هستند: برخی توانایی حرکت دارند و برخی توانایی حرکت ندارند. در مجرای اسپرمبر اسپرمهای سالم قطعا توانایی حرکت دارند.
مجرای اسپرمبر و اپیدیدیم طویل هستند، اما مجرای اسپرمساز طویل نیست.
تمامی مجاری اسپرمساز به اپیدیدیم راه دارند.
بخشی از اپیدیدیم که در تماس با اسپرمساز است از بخشی که با اسپرمبر در تماس نیست قطورتر است.
در دیواره لوله اسپرمساز، خارجیترین تقسیم تقسیم میتوز و داخلیترین تقسیم تقسیم میوز ۲ است.
هر سلول هاپلوئید در مرد حاصل از تقسیم نیست.
اسپرماتوگونیها همانند اسپرماتوسیتهای ثانویه به هم متصل هستند.
ژن بیان تاژک در اسپرماتید بیان میشود. اسپرماتید میتواند تاژک داشته باشد.
ژن بیان تاژک روی کروموزومهای غیرجنسی قرار دارد.
در زمان تمایز اسپرماتوسیت ثانویه هسته فشرده میشود.
در زمان تمایز اسپرماتید هسته فشرده میشود نه کروموزومهام. اسپرمها و اسپرماتیدها در G0 قرار دارند.
در تمایز اسپرماتید چنین اتفاقی میافتد: سلولها از هم جدا و تاژکدار میشود، سپس مقداری از سیتوپلاسم خود را از دست میدهد، هسته آن فشرده شده و در سر به صورت مجزا قرار میگیرد و سلول حالت کشیده پیدا میکند.
اولین بخشی از اسپرماتید که از لوله اسپرمساز خارج میشود تاژک آن است.
سلول سرتولی با ترشحات خود تمایز اسپرمها را هدایت میکند.
به دست آوردن توانایی حرکت هم خود نوعی تمایز است. درواقع محل بلوغ اسپرمها در اپیدیدیم است.
تا قبل از وزیکول سمینال غذا اسپرم را سلولهای سرتولی تامین میکند.
سلول سرتولی و برخی از نوروگلیاها جز دستگاه ایمنی نیستند، اما بیگانهخواری را انجام میدهند.
در سر و تنه اسپرم تعداد زیادی DNA دیده میشود. در سر هسته وجود دارد و در تنه تعداد زیادی میتوکندری.
انتهای دم اسپرم بدون پوشش است.
غدد ضمیمهای دستگاه تولید مثل ۵تاست و۳ نوع: دو تا پیاز میزراهی، دو تا وزیکول سمینال و یک پروستات.
پروستات برخلاف پیاز میزراهی در مجاورت مثانه قرار دارد.
وزیکول سمینال پشت مثانه قرار دارد، پروستات و پیاز میزراهی زیر مثانه قرار دارد.
ترشحات وزیکول سمینا ابتدا وارد مجرای خود و سپس وارد مجرای اسپرمبر میشود.
وزیکول سمینال فقط فروکتوز ندارد. این غده مایع غنی از فروکتوز دارد.
ترشحات پروستات شیریرنگ و قلیاییست.
ترشحات پیاز میزراهی روانکننده و قلیاییست.
ترشحات قلیایی مسیر حرکت اسپرمها را خنثی میکند.
اسپرم بخشی از مایع منی نیست.
دو مجرای اسپرمبر از زیر مثانه وارد پروستات میشود.
مجرای اسپرمبر از جلوی میزنای و پشت مثانه عبور میکند.
در تولیدمثل جنسی، دو یاخته جنسی(گامت) باهم ترکیب و هستههای آنها باهم ادغام میشوند. یاختههای موثر در تولیدمثل جنسی با نوعی تقسیم کاهشی به نام کاستمان ایجاد میشوند. به نظر شما اهمیت این نوع تقسیم در جانداران چیست؟
کاستمان از دو مرحله کلی کاستمان ۱ و ۲ تشکیل شده است؛ پس از تقسیم هسته نیز تقسیم سیتوپلاسم انجام میشود. پیش از این تقسیم نیز، مانند رشتمان، اینترفاز رخ میدهد.
کاستمان ۱
در این مرحله از تقسیم کاستمان، عدد فامتنی نصف میشود. این بخش از کاستمان چهار مرحله دارد که عبارتاند از: پروفاز ۱، متافاز۱، آنافاز۱ و تلوفاز۱.
پروفاز ۱: فامتنهای همتا از طول در کنار هم قرار میگیرند و فشرده میشوند. به این ساختار چهارفامینکی، چهارتایه(تتراد) گفته میشود. چهارتایه از ناحیه سانترومر به رشتههای دوک متصل میشوند. سایر وقایع این مرحله، شبیه پروفاز و پرومتافاز رشتمان است.
متافاز ۱: چهارتایهها در استوای یاخته، روی رشتههای دوک قرار میگیرند.
آنافاز ۱: فامتنهای همتا که مضاعف شدهاند، از هم جدا میشوند و به سمت قطبین یاخته حرکت میکنند. نحوه کوتاهشدن رشتههای دوک، شبیه فرایند رشتمان است.
تلوفاز ۱: با رسیدن فامتنها به دو سوی یاخته، پوشش هسته دوباره تشکیل میشود. معمولا درپایان کاستمان ۱ تقسیم سیتوپلاسم انجام میشود. نتیجهی کاستمان ۱ ایجاد دویاخته است.
کاستمان ۲
در این مرحله یاختههای حاصل از کاستمان ۱، مراحل پروفاز۲، متافاز۲، آنافاز۲ و تلوفاز ۲ رامیگذرانند. وقایع کاستمان ۲ بسیار شبیه رشتمان است و در پایان آن، از هر یاخته دو یاخته شبیه هم ایجاد میشود که نصف فامتنهای یاختههای مادر را دارند. این فامتنها مضاعف نیستند. در پایان کاستمان ۲، تقسیم سیتوپلاسم انجام میشود. در مجموع و با پایان تقسیم کاستمان از یک یاخته ۲n، چهار یاخته n فامتنی حاصل میشود.
تغییر در تعداد فامتنها
گرچه تقسیم یاختهای با دقت زیاد انجام میشود، ولی به ندرت ممکن است اشتباهاتی در روند تقسیم رخ دهد. چندلادی (پلیپلوئیدی) شدن و با هم ماندن فامتنها، نمونههایی از این خطاهای کاستمانی هستند. اشتباه در تقسیم میتواند، هم در تقسیم رشتمان و هم در تقسیم کاستمان رخ دهد، ولی چون یاختههای حاصل از کاستمان در ایجاد نسل بعد دخالت مستقیم دارند، از اهمیت بیشتری برخوردارند.
چندلادی شدن: اگر در مرحله آنافاز همه فامتنها بدون اینکه از هم جدا شوند به یک یاخته بروند، آن یاخته دو برابر فامتن خواهد داشت و یاخته دیگر فاقد فامتن خواهد بود. در آزمایشگاه میتوان با تخریب رشتههای دوک تقسیم این وضعیت را ایجاد کرد.
به یاخته یا جانداری که یاختههای آن بیش از دو مجموعه فامتن داشته باشد، چندلاد گفته میشود؛ مثلًا گندم زراعی sn و موز 3n است)شکل .(17
با هم ماندن فامتنها: در این حالت، یک یا چند فامتن در مرحله آنافاز(رشتمان و کاستمان) از هم جدا نمیشوند. بنابراین، در یاختههای حاصل، کاهش یا افزایش یک یا چند فامتن مشاهده میشود. نمونه این حالت، نشانگان داون است. به آمیزهای از نشانههای یک بیماری، یا یک حالت نشانگان میگویند. افراد مبتلا به داون، در یاختههای پیکری خود 47 فامتن دارند. فامتن اضافی مربوط به شماره 21 است؛ یعنی یاختههای پیکری این افراد ۳ فامتن شماره ۲۱ دارند. علت بروز این حالت آن است که یکی از یاختههای جنسیِ ایجادکننده فرد، بهجای یک فامتن شماره ۲۱، دارای دو فامتن ۲۱ بوده است. بالابودن سن مادران در هنگام بارداری از عوامل مهم بروز این بیماری است؛ زیرا با افزایش سن مادر، احتمال خطای کاستمانی در تشکیل یاختههای جنسی وی بیشتر میشود. علت این موضوع را در فصلهای آینده خواهید آموخت.
عوامل محیطی نیز میتوانند موجب اختلال در تقسیم کاستمان شوند. دخانیات، الکل، مجاورت با پرتوهای مضر و آلودگیها نیز میتوانند در روند جدا شدن فامتنها در هر دو جنس، اختلال ایجاد کنند.
تولید مثل جنسی لزوما در اثر لقاح رخ نمیدهد.
تولید مثل جنسی میتواند با حضور یک یا دو والد رخ دهد.
تنها شرط تولید مثل جنسی حضور گامت است.
شرط تشکیل تتراد کنار هم قرار گرفتن کروموزومهای همتاست. درواقع زمانی که کروموزموم X و Y در کنار هم قرار میگیرد ساختار تتراد تشکیل نمیشود.
در پروفاز یک تتراد تشکیل و رشته دوک به سانترومر متصل میشود. در پروفاز یک برخلاف میتوز به هر سانترومر یک رشته دوک متصل میشود.
در آنافاز یک برخلاف میتوز تعداد کروموزومها ثابت میماند.
در مراحل مختلف میوز برخلاف میتوز هیچ کشیدگیای در سلول رخ نمیدهد.
در فاصله میوز یک و دو اندازه کروموزوم تغییر خاصی نمیکند.
در فاصله میوز یک و دو برخی اندامکها مثل سانتریول و میتوکندری زیاد میشود. این یعنی سلول مرحله G2 را رد میکند.
اووسیت ثانویه در حدفاصل میوز یک و دو متوقف میشود.
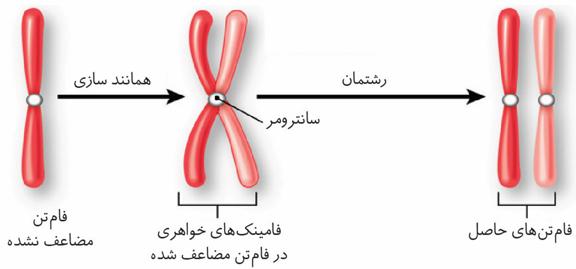
دررِشتِمان ماده ژنتیک،که در مرحله S همانندسازی شده بود، تقسیم میشود و به یاختههای جدید میرسد. فامتنها که در هسته پراکندهاند، ابتدا باید بهطور دقیق در وسط یاخته آرایش یابند و به مقدار مساوی بین یاختههای حاصل تقسیم شوند.
برای حرکت و جداشدن صحیح فامتنها، ساختارهایی به نام دوک تقسیم ایجاد میشود. دوک تقسیم، مجموعهای از ریزلولههای پروتئینی است که هنگام تقسیم، پدیدار و سانترومر فامتن به آن متصل میشود.
با کوتاه شدن رشتههای دوک متصل به سانترومر، فامتنها ازهم جدا میشوند و به قطبین میروند.
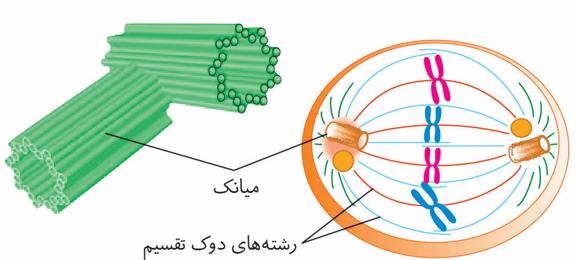
در یاختههای جانوری، میانکها(سانتریولها) ساخته شدن رشتههای دوک را سازمان میدهند.
هر میانک ساختاری استوانهای شکل است. در یاخته دو عدد میانک بهصورت عمود بر هم وجود دارند که در اینترفاز، برای تقسیم یاخته، دوبرابر میشوند. هر میانک، از نُه دسته سهتایی از ریزلولههای پروتئینی تشکیل شده است.
سانتریولها جز اندامکهای بدون غشا محسوب میشوند.
بررسی کنید
در هر یاختهی در حال میتوز قطعا یک سر رشته دوک به سانتریول متصل میشود؟
پاسخ
سلولهای گیاهی تقسیم میتوز انجام میدهند، اما سازماندهی رشتههای دوک در آنها توسط سانتریولها انجام نمیشود. حالا بیایید سوال دیگری را بررسی کنیم:
در هر یاخته جانوری در حال میتوز، قطعا یک سر رشته دوک به سانتریول متصل میشود؟
در سطح کتاب درسی ما سه نوع رشته دوک داریم: رشتههایی که به وسط سلول نمیآیند. رشتههایی که به وسط سلول میآیند اما به سانترومر متصل نمیشوند و رشتههایی که تا وسط سلول میآیند و به سانترومر متصل میشوند.
همه این رشتهها به سانتریول متصل میشوند.
تقسیم هسته
رشتمان، فرایندی پیوسته است، ولی زیستشناسان برای سادگی، آن را مرحلهبندی میکنند.
پروفاز:
در این مرحله، رشتههای فامینه فشرده، ضخیم و کوتاهتر میشوند. بهطوریکه بهتدریج با میکروسکوپ نوری میتوان آنها را مشاهده کرد. ضمن فشرده شدن فامتن، میانکها به دو طرف یاخته حرکت میکنند و بین آنها دوک تقسیم تشکیل میشود. در این مرحله پوشش هسته شروع به تخریب میکند.
در پروفاز فشرده شدن کروموزومها مقدم بر دور شدن سانتریولهاست.
پرومتافاز:
در این مرحله، پوشش هسته و شبکه آندوپلاسمی تجزیه میشوند تا رشتههای دوک بتوانند به فامتنها برسند. درهمین حال سانترومر فامتنها به رشتههای دوک متصل میشوند.
متافاز:
فامتنها بیشترین فشردگی را پیدا میکنند و در وسط (سطح استوایی) یاخته ردیف میشوند.
آنافاز:
در این مرحله، با تجزیه پروتئین اتصالی در ناحیه سانترومر، فامینکها از هم جدا میشوند. فاصله گرفتن فامینکها با کوتاه شدن رشتههای دوک متصل به فامتن انجام میشود. فامتنها که اکنون تکفامینکیاند، به دو سوی یاخته (قطب) کشیده میشوند.
تلوفاز:
رشتههای دوک تخریب شده و فامتنها شروع به باز شدن میکنند تا به صورت فامینه درآیند. پوشش هسته نیز مجدداً تشکیل میشود. در پایان تلوفاز، یاخته، دو هسته مشابه دارد.
تقسیم سیتوپلاسم
پس از رشتمان، اجزای یاخته بین دو سیتوپلاسم تقسیم میشوند. با تقسیم سیتوپلاسم دو یاختۀ جدید تشکیل میشود.
تقسیم در سلولهای جانوری
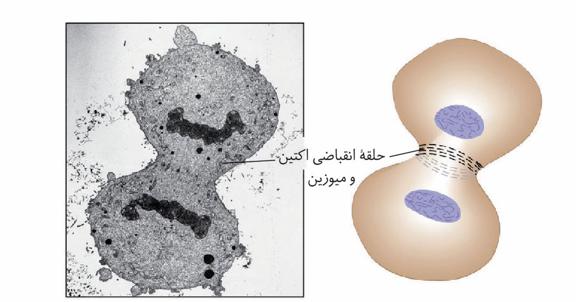
در یاختههای جانوری تقسیم سیتوپلاسم با ایجاد فرورفتگی در یاخته شروع میشود. این فرورفتگی حاصل انقباض حلقهای از جنس اکتین و میوزین است که مانند کمربندی در سیتوپلاسم قرار میگیرد و به غشا متصل است. با تنگ شدن این حلقه انقباضی در نهایت دو یاخته از هم جدا میشوند.
تقسیم در سلولهای گیاهی
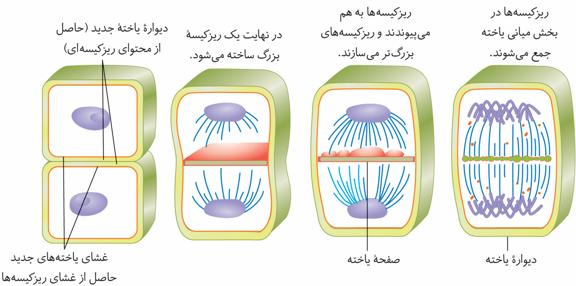
در یاختههای گیاهی، حلقه انقباضی تشکیل نمیشود. در این یاختهها نخست ساختاری به نام صفحه یاختهای در محل تشکیل دیواره جدید، ایجاد میشود. این صفحه با تجمع ریزکیسههایدستگاه گلژی و بههم پیوستن آنها تشکیل میشود. این ریزکیسهها، دارای پیشسازهای تیغه میانی و دیواره یاختهاند. با اتصال این صفحه به دیواره یاخته مادری دو یاخته جدید از هم جدا میشوند. ساختارهایی مانند لان و پلاسمودسم در هنگام تشکیل دیواره جدید، پایهگذاری میشوند.
در یاختههای گیاهی سازماندهی رشتههای دوک انجام میشود، اما این سازماندهی توسط سانتریول نیست.
تقسیم سلولی فرایندی تنظیم شده است
بعضی یاختههای بدن جانداران، مانند یاختههای بنیادی مغز استخوان و یاختههای مریستمی گیاهان میتوانند دائما تقسیم شوند. همین یاختهها در شرایطی خاص، مثلا شرایط نامساعد محیطی یا افزایش بیش از حد تعداد یاختهها، تقسیم خود را کاهش میدهند و یا متوقف میکنند.
برعکس، یاختههای عصبی به ندرت تقسیم میشوند. این یاختهها چگونه تشخیص میدهند در چه زمان یا به چه مقداری باید تقسیم شوند؟ چه عواملی تنظیمکننده سرعت و تعداد تقسیم یاختهاند؟ چگونه تعداد چرخههای یاخته تنظیم میشوند و چرا این تنظیم دربرخی یاختهها به هم میخورد؟
عوامل تنظیم کننده تقسیم سلول
یاختهها در پاسخ به بعضی عوامل محیطی و مواد شیمیایی سرعت تقسیم خود را تنظیم میکنند.
انواعی از پروتئینها وجود دارد که با فرایندهایی منجر به تقسیم یاختهای میشوند. پروتئینهای دیگری نیز وجود دارند که در شرایط خاصی، مانع از تقسیم یاختهها میشوند.
این پروتئینها در سرعت تقسیم یاخته مانند پدال گاز و ترمز عمل میکنند؛ یا در گیاهان در محل آسیبدیده، نوعی عامل رشد تولید میشوند تا با تقسیم سریع، توده یاخته ایجاد کنند. این توده یاخته مانع نفوذ میکروبها میشود؛ یا نوعی عامل رشد، در پوست انسان زیرمحل زخم تولید میشود که با افزایش سرعت تقسیم یاختهها، سرعت بهبود زخم را افزایش میدهد.
مثال دیگر این مواد، اریتروپویتین است. این ماده بر کدام بخش بدن اثر میگذارد و نتیجه آن چیست؟
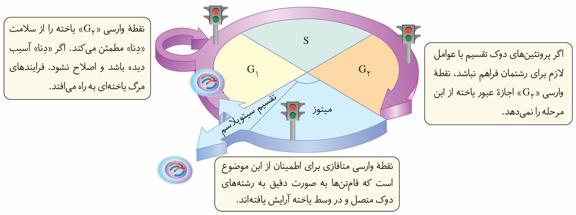
در چرخه یاختهای، چند نقطه وارسی وجود دارد. نقاط وارسی، نقاطی از چرخه یاختهاند که به آن اطمینان میدهند که مرحله قبل کامل شده است و عوامل لازم برای مرحله بعد آمادهاند.
نقطه وارسی G۱
نقطه وارسی «G۱» یاخته را از سلامت «دِنا» مطمئن میکند. اگر «دِنا» آسیب دیده باشد و اصلاح نشود، فرایندهای مرگ یاختهای بهراه میافتد.
نقطه وارسی G۲
اگر پروتئینهای دوک تقسیم یا عوامل لازم برای رشتمان فراهم نباشد، نقطه وارسی «G۲» اجازه عبور یاخته از این مرحله را نمیدهد.
نقطه وارسی متافازی
نقطه وارسی متافازی برای اطمینان از این موضوع است که فامتنها بهصورت دقیق به رشتههای دوک متصل و در وسط یاخته آرایش یافتهاند.
تقسیم بیرویه سلول
یاختهها با تقسیم، افزایش و با مرگ، کاهش مییابند. اگر تعادل بین تقسیم یاخته و مرگ یاختهها بههم بخورد، چه وضعی پیش میآید؟ نتیجه میتواند ایجاد یک تومور باشد. تومور، تودهای است که در اثر تقسیمات تنظیم نشده ایجاد میشود.
تومورها به دو نوع خوشخیم و بدخیم تقسیم میشوند.
تومور خوشخیم
نوع خوشخیم رشدی کم دارد و یاختههای آن در جای خود میمانند و منتشر نمیشوند. این نوع تومور معمولا آنقدر بزرگ نمیشود که به بافتهای مجاور خود آسیب بزند. البته در مواردی که تومور بیش از اندازه بزرگ شود، میتواند در انجام اعمال طبیعی اندام اختلال ایجاد کند.
لیپوما یکی از انواع تومورهای خوشخیم است که در افراد بالغ متداول است. در این تومور، یاختههای چربی تکثیر شده و توده یاخته ایجاد میکند.
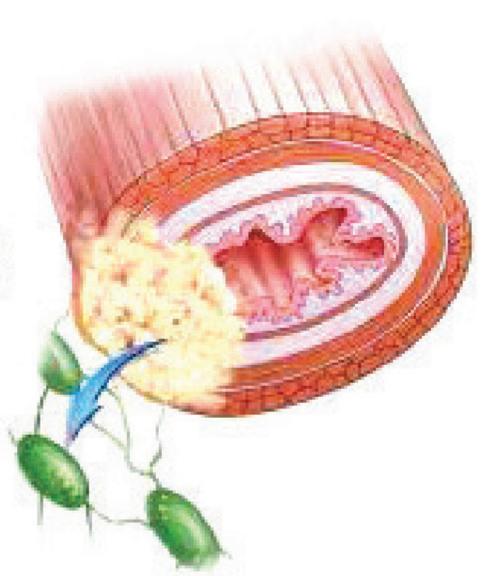
تومور بدخیم(سرطان)
تومور بدخیم یا سرطان به بافتهای مجاور حمله میکند؛ یاختههایی از این تومورها میتوانند جدا شوند و همراه با جریان خون، یا به ویژه لنف به نواحی دیگر بدن بروند، در آنجا مستقر شوند و رشد کنند(علت اصلی سرطان، بعضی تغییرات در مادۀ ژنتیکی یاخته است که باعث میشود چرخه یاخته از کنترل خارج شود).
مراحل پیشرفت تومور بدخیم
مرحله یک: یاخته سرطانی شروع به تهاجم به یاختههای بافت میکند.
مرحله دو: یاختههای سرطانی در بافتها گسترش مییابند، ولی هنوز به دستگاه لنفی مجاور راه پیدا نکردهاند.
مرحله سه: یاختههای سرطانی به بخشهای لنفی مجاور محل تکثیر خود، دسترسی پیدا میکنند.
مرحله چهار: یاختههای سرطانی از راه لنف به بافتهای دورتر میروند و پس از استقرار موجب سرطانی شدن آنها میشوند.
تشخیص و درمان سرطان
روشهای متعددی برای تشخیص و درمان سرطانها وجود دارد و گاهی ترکیبی از این روشها مورد استفاده قرار میگیرد. روشهای رایج درمان سرطان شامل جراحی، شیمیدرمانی و پرتودرمانی است.
بافتبرداری روشی است که در آن، تمام یا بخشی از بافت سرطانی یا مشکوک به سرطان برداشته میشود. آزمایش خون به این شناسایی کمک میکند.
در پرتودرمانی، یاختههایی که به سرعت تقسیم میشوند، بهطور مستقیم تحتتاثیر پرتوهای قوی قرار میگیرند.
شیمی درمانی با استفاده از داروها باعث سرکوب تقسیم یاختهها در همه بدن میشود. این روشهای درمانی میتوانند به یاختههای مغز استخوان، پیاز مو و پوشش دستگاه گوارش نیز آسیب برسانند. مرگ این یاختهها از عوارض جانبی شیمیدرمانی است که باعث ریزش مو، تهوع و خستگی میشود. حتی بعضی افراد که تحتتاثیر تابشهای شدید یا شیمیدرمانی قوی قرار میگیرند مجبور به پیوند مغز استخوان میشوند تا بتوانند یاختههای خونی مورد نیاز را بسازند.
وراثت و محیط، هر دو در ایجاد سرطان موثر هستند
پروتئینها، تنظیمکننده چرخه یاخته و مرگ آن هستند. پروتئینها محصول عملکرد ژنها هستند. بنابراین، مشخص است که در ایجاد سرطان، ژنها نقش دارند. ژنهای زیادی شناخته شدهاند که در بروز سرطان موثرند. علت شیوع بیشتر بعضی سرطانها در بعضی جوامع، همین مسئله است.
عوامل محیطی هم در بروز سرطان موثرند. پرتوهای فرابنفش، بعضی آلایندههای محیطی و دود خودروها به ساختار «دِنا» آسیب میزنند. سایر پرتوها و مواد شیمیایی سرطانزا، موادغذایی دودی شده مثل گوشت و ماهی دودی، بعضی ویروسها، قرصهای ضدبارداری، نوشیدنیهای الکلی و دخانیات از عوامل مهم سرطانزاییاند.
مرگ برنامهریزی شده
مرگ یاختهها میتواند تصادفی باشد؛ مثلًا در بریدگی، یاختهها آسیب میبینند و از بین میروند. به این حالت، بافت مردگی گفته میشود. ولی مرگ برنامهریزی شده یاختهای شامل یک سری فرایندهای دقیقا برنامهریزی شده است که در بعضی یاختهها و در شرایط خاص ایجاد میشود. این فرایند با رسیدن علایمی به یاخته شروع میشود. به دنبال این رخداد، در چند ثانیه پروتئینهای تخریبکننده در یاخته شروع به تجزیه اجزای یاخته و مرگ آن میکنند.
حذف یاختههای پیر یا آسیبدیده، مانند آنچه در آفتابسوختگی اتفاق میافتد، مثالی از مرگ برنامهریزی شده یاختهای است؛ چون پرتوهای خورشید دارای اشعه فرابنفشاند آفتابسوختگی میتواند سبب آسیب به «دِنا» یاختهها و بروز سرطان شود. مرگ برنامهریزی شده یاختهای، با از بین بردن یاختههای آسیبدیده، آنها را حذف میکند. مثال دیگر، حذف یاختههای اضافی از بخشهای عملکردی مانند پردههای بین انگشتان پا در پرندگان است.
در اینترفاز ممکن است رشتههای میکروتوبولی دیده شود، اما دقت کنید این رشتههای میکروتوبولی دوک تقسیم نیستند.
مراحل تقسیم
در پروفاز غشای هسته شروع به تخریب میکند، اما در پرومتافاز به طور کامل تجزیه میشود.
پرومتافاز اولین مرحلهایست که کروموزومها در تماس با سیتوپلاسم قرار میگیرند.
در متافاز هسته وجود ندارد و سلولها در سطح استوایی یاخته (نه هسته) ردیف میشوند.
در پرومتافاز شبکه آندوپلاسمی زبر تجزیه میشود.
در تلوفاز شبکه آندوپلاسمی زبر دوباره تشکیل میشود.
اولین مرحلهای که رشتههای دوک به سانترومر متصل میشود پرومتافاز است.
در متافاز کروموزوم به حداکثر فشردگی خود میرسد. این فشردگی در آنافاز هم ادامه دارد. درواقع در متافاز و آنافاز حداکثر فشردگی در کروموزوم دیده میشود. در تلوفاز کروموزومها شروع به باز شدن میکنند.
در متافاز برای حرکت دادن کروموزومها برخی از رشتههای دوک تقسیم بلند و برخی دیگر کوتاه میشوند. در آنافاز هم کوتاه شدن رشتههای دوک تقسیم دیده میشود.
در آنافاز سلول عرض سلول بیشتر میشود.
در آنافاز کروموزومهای یاخته دوبرابر میشود.
سیتوکینز الزامی نیست و ممکن است گاهی رخ ندهد، اما اگر رخ داد نکات زیر در مورد آن صادق است:
سیتوکینز در سلول جانوری در حین تلوفاز شروع میشود.
سیتوکینز در سلول گیاهی از اواخر آنافاز شروع میشود و در تمام تلوفاز ادامه دارد.
در سلول گیاهی رشتهّای دوکی وجود دارد که ریزکیسههای غشایی را به صفحه یاختهای هدایت میکند.
در یاختههای جانوری سیتوکینز با فرورفتگی غشایی آغاز میشود. دقت کنید این فرورفتگی الزاما در وسط سلول نیست.
کمربندی که باعث فرورفتگی غشایی میشود به درون غشا متصل میشود نه بیرون از آن.
این کمربند از جنس اکتین و میوزین است. پس هر سلولی که اکتین و میوزین در حال فعالیت هستند سلول ماهیچهای نیست. در واقع هر سلولی که روی تقسیم اثرگذار هستند میتوانند روی فعالیت اکتین و میوزین اثر بگذارد.
دقت کنید که ما در بحث تقسیم سلول جانوری حلقهی انقباضی داریم و اکتین و میوزین منقبض نمیشوند.
سیتوکینز نامساوی با سیتوکینز ناکامل فرق میکند. نتیجه سیتوکینز نامساوی دو سلول است که هم اندازه نیستند و نتیجه سیتوکینز ناکامل سلولهای به هم متصل است.
در سیتوکینز سلول گیاهی تیغه میانی تشکیل میشود. یعنی ریزکیسهها حاوی موادی هستند که تیغه میانی را تشکیل میدهد. دقت کنید این ریزکیسهها دیواره نخستین را نمیسازند چون دیوار نخستین توسط سلولهای تازه تشکیل ساخته میشود.
لان و پلاسمودسم در هنگام تشکیل دیواره پایه گذاری میشود (عجب جمله مسخرهای).
هم در G1 و هم در G2 میتوان اصلاح DNA خطی را دید.
سلولهای تومور خوش خیم سرطانی نمیشوند، اما ممکن است آلوده به ویروس شده و مورد حمله لنفوسیتهای T قرار بگیرد.
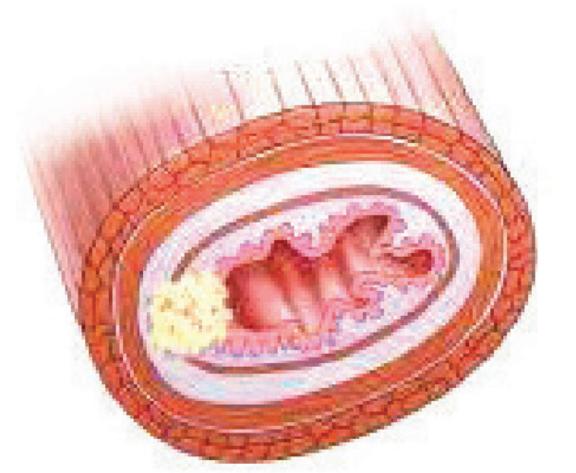
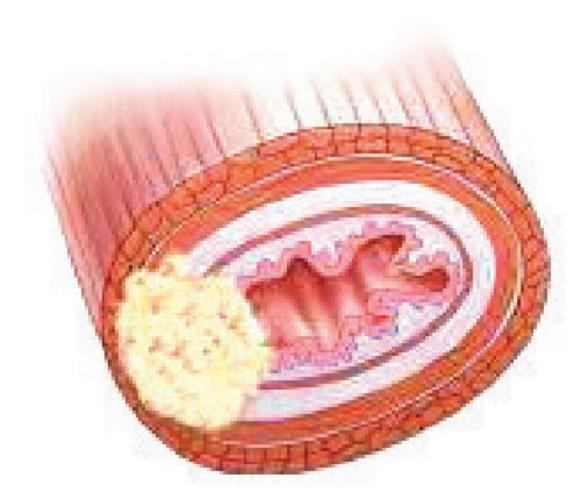
مراحل سرطانی شدن لوله گوارش (ترتیب شمارهها همان ترتیب عکسها در کتاب است).
۱- درگیر شدن مخاط و زیر مخاط
۲-درگیر شدن لایه ماهیچهای
۳-درگیر شدن لایه بیرونی
آپوپتوز در کتاب درسی:
سلولهای سالمی که آپوپتوز میشوند: پیر – پیوندی – سلولهای اضافی که در روند تکوین حذف میشود.
سلولهای ناسالمی که آپوپتوز میشوند: سلول آلوده به ویروس – سلول سرطانی – سلول آسیب دیده